
文章阅读目录大纲
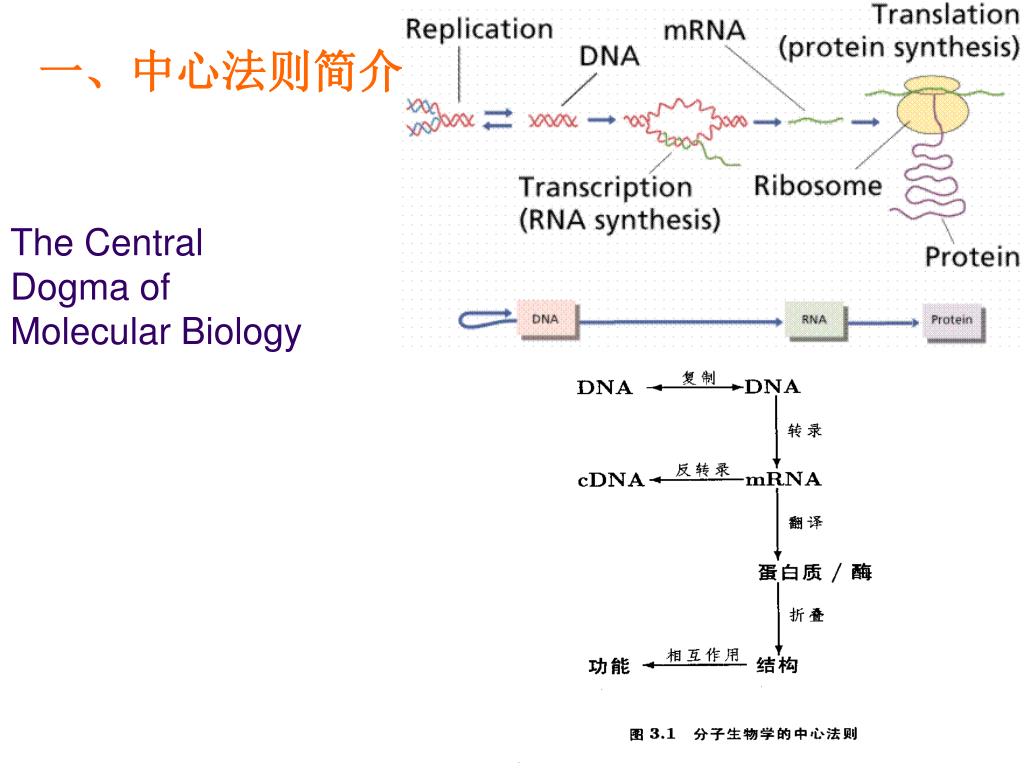
原核生物细胞内的中心法则是指遗传信息从DNA经RNA到蛋白质的传递过程,具有高效和经济的特点。DNA复制、转录和翻译均在细胞质中进行,且转录与翻译高度偶联——新生mRNA尚未完全合成,核糖体便已结合并开始翻译,极大提升了蛋白质合成速率。原核生物mRNA常为多顺反子结构,一条mRNA可编码多个功能相关的蛋白质,且无内含子、无需剪接,可直接作为翻译模板。此外,原核mRNA半衰期极短,便于快速响应环境变化。基因表达主要通过操纵子结构在转录水平进行精细调控,如乳糖操纵子和色氨酸操纵子,使原核生物能够灵活适应多变环境。这些机制共同构成了原核生物中心法则的核心,体现了其高度优化的遗传信息传递系统。
将复杂的生物学过程转化为可计算的数学模型是系统生物学的核心。最近的工作中需要将原本非常粗糙的虚拟细胞翻译事件模型拆解为更加细分化的多步骤生物化学过程,以适应针对细胞群落生长的建模计算。原核生物的翻译过程一般可以拆解为四个主要阶段:起始、延伸、终止和再循环。下面为我将原核生物的翻译过程拆解为一系列可以用化学式表示的单元步骤的结果。
在介绍这些分步骤之前,为了方便书写和理解,我们会需要首先来定义一下模型中会用到的各种“化学物质”(分子和复合物):
- 核糖体: R_30S (小亚基), R_50S (大亚基), R_70S (完整核糖体)
- mRNA: mRNA
- tRNA: tRNA^aa (携带氨基酸aa的tRNA)
- 氨基酸: aa (泛指), fMet (甲酰甲硫氨酸)
- 氨酰-tRNA: aa-tRNA^aa (已“充电”的tRNA)
- 多肽链: P_n (含有n个氨基酸的多肽链)
- 能量分子: GTP, GDP, Pi (无机磷酸), ATP, AMP, PPi (焦磷酸)
- 起始因子: IF1, IF2, IF3
- 延伸因子: EF-Tu, EF-Ts, EF-G
- 释放因子: RF1, RF2, RF3
- 酶: aaRS (氨酰-tRNA合成酶)
关于翻译所需的核糖体复合物的描述
Prokka 是一个广泛使用的原核生物基因组注释工具。它会通过搜索保守的 RNA 基因数据库(如 RFAM)来识别基因组中的 rRNA 和 tRNA 基因。对于细菌和古菌,其核糖体 RNA 的标准构成就是 5S、16S 和 23S rRNA,因此在原核基因组中,假若我们通过prokka软件进行基因注释,一般会从完整的基因组序列中注释出5s, 16s, 23s这三种核糖体RNA的基因。对于在这里进行翻译所需要的30s核糖体亚基,50s核糖体亚基和完整的70s核糖体亚基,他们之间所构成的复合体的组装关系一般为:
5s + 23s = 50s
16s = 30s
50s(大亚基) + 30s(小亚基) = 70s核糖体的组装是一个被称为“核糖体生物发生”的极其复杂、高度有序的过程,涉及 rRNA 的转录、加工、折叠,以及核糖体蛋白按特定顺序的结合,还需要许多组装因子和分子伴侣的辅助。上面所给到的关系是一种简化后的表示方法,在实际的细胞环境中,更加复杂准确的描述可以为:
- 30S 小亚基的构成:由 1 分子 16S rRNA 和 约 21 种核糖体蛋白(Ribosomal Proteins, 命名为 S1, S2, S3…S21)共同构成。
- 50S 大亚基的构成:由 1 分子 23S rRNA、1 分子 5S rRNA 和 约 34 种核糖体蛋白(命名为 L1, L2, L3…L36)共同构成。
- 70S 核糖体的组装:成熟的 30S 小亚基 和 50S 大亚基 在 mRNA 和起始因子(Initiation Factors, IF1, IF2, IF3)的介导下,结合形成有功能的 70S 起始复合体,准备开始翻译。 50S + 30S = 70S 描述的是功能性复合体的最终形成步骤,这个过程本身也需要其他因子的参与。
对于核糖体亚基上的数字,其一般表示核糖体这些生物分子的在进行离心时的沉降速率而定的。测量沉降速率的单位是Svedberg(S,S值越大,沉降速率越快),Svedberg (S) 是一个时间单位 (1 S = 10⁻¹³ 秒),它表示颗粒在超速离心场中沉降的速率,并且S值不具有加和性。当两个亚基结合时,它们的总表面积减小,形状变得更加紧凑,导致整体的流体阻力特性发生改变,因此沉降速率(S值)并非简单相加。上面的这些数字并不直接等同于字面上的质量。这就是为什么 50S 亚基和 30S 亚基结合后,形成的 70S 核糖体的 S 值 不是 50 + 30 = 80 S。70S 小于 80S,正是因为结合后的复合体形状和结构发生了变化。
在原核细胞内,蛋白质翻译是一个高效而精确的过程,通常被分为四个主要阶段。下面我们用一个简单的比喻来理解:把核糖体想象成一个蛋白质工厂,mRNA是生产蓝图,tRNA是运送特定零件(氨基酸)的卡车。以下为对所涉及到的4个阶段的介绍。
阶段一:翻译起始
此阶段的目标是组装一个功能性的70S起始复合物,其中核糖体的小亚基已结合到mRNA的起始密码子上,起始tRNA(携带甲酰甲硫氨酸,fMet)已就位于P位点。
单元步骤 1:30S小亚基与mRNA的结合
30S核糖体亚基在起始因子IF1和IF3的帮助下,识别并结合到mRNA的Shine-Dalgarno(SD)序列上,形成30S前起始复合物。在这个过程中会涉及到一些关键的分子复合物,例如30S亚基, mRNA, 起始因子IF1, 起始因子IF3。需要注意的是,这个步骤一个可逆的结合反应,但是IF3的存在会阻止50S亚基过早结合,确保了反应的有序性。
30S + mRNA + IF1 + IF3 <=> 30S_mRNA_IF1_IF3单元步骤 2:起始tRNA的装载
携带甲酰甲硫氨酸的起始tRNA(fMet-tRNA^fMet)与GTP结合的起始因子IF2形成复合物,然后进入30S前起始复合物的P位点。在这里,同样是可逆结合反应。IF2-GTP对fMet-tRNA^fMet具有特异性,确保了正确的起始tRNA被招募。在这里会涉及到下面的三种关键的复合物因子:fMet-tRNA^fMet, 起始因子IF2, GTP。
fMet-tRNA^fMet + IF2-GTP <=> fMet-tRNA^fMet_IF2-GTP
30S_mRNA_IF1_IF3 + fMet-tRNA^fMet_IF2-GTP <=> 30S_mRNA_fMet-tRNA^fMet_IF1_IF2-GTP_IF3单元步骤 3:50S大亚基的结合与起始因子的释放
这是一个由GTP水解驱动的、基本不可逆的反应。在模型中,正向反应速率应远大于逆向速率。50S大亚基结合到上述复合物上,形成完整的70S起始复合物。此过程触发IF2上的GTP水解为GDP和Pi,释放的能量导致所有起始因子(IF1, IF2-GDP, IF3)解离。
30S_mRNA_fMet-tRNA^fMet_IF1_IF2-GTP_IF3 + 50S -> 70S_mRNA_fMet-tRNA^fMet + IF1 + IF2-GDP + IF3 + Pi阶段二:翻译延伸
此阶段是一个循环过程,不断将氨基酸添加到正在生长的多肽链上。每个循环包括三个主要步骤。
单元步骤 1:氨酰-tRNA的进位
一个与延伸因子EF-Tu和GTP结合的氨酰-tRNA进入核糖体的A位点。这个过程中会涉及到下面的几个关键的复合物:氨酰-tRNA (aa-tRNA), 延伸因子EF-Tu, GTP。基于上面的几个复合物,EF-Tu-GTP作为aa-tRNA的递送载体。当正确的aa-tRNA(其反密码子与A位点的mRNA密码子配对)进入后,GTP被水解,EF-Tu-GDP释放。这是一个关键的校对步骤,确保翻译的保真性。
aa-tRNA + EF-Tu-GTP <=> aa-tRNA_EF-Tu-GTP
70S_mRNA_pept-tRNA(P-site) + aa-tRNA_EF-Tu-GTP -> 70S_mRNA_pept-tRNA(P-site)_aa-tRNA(A-site) + EF-Tu-GDP单元步骤 2:肽键形成
核糖体大亚基70S核糖体 (肽酰转移酶活性)的肽酰转移酶中心催化P位点上肽酰-tRNA的肽链转移到A位点的氨酰-tRNA的氨基上,形成一个新的肽键。这是翻译的核心化学反应。其速率通常非常快,可以用一个催化速率常数 (k_cat) 来描述。该步骤不消耗额外的能量(如ATP/GTP),能量已在tRNA装载和进位步骤中消耗。
70S_mRNA_pept-tRNA(P-site)_aa-tRNA(A-site) -> 70S_mRNA_deacyl-tRNA(P-site)_pept-aa-tRNA(A-site)单元步骤 3:转位
核糖体沿着mRNA移动一个密码子的距离。A位点上的肽酰-tRNA移动到P位点,P位点上脱酰化的tRNA移动到E位点并离开。此过程由延伸因子EF-G和GTP水解驱动的不可逆步骤。它确保了翻译过程的单向性。EF-G-GTP的结合和水解是模型的关键动力学事件。
70S_mRNA_deacyl-tRNA(P-site)_pept-aa-tRNA(A-site) + EF-G-GTP -> 70S_mRNA_pept-aa-tRNA(P-site) + EF-G-GDP + Pi + deacyl-tRNA(E-site)阶段三:翻译终止
当核糖体的A位点遇到一个终止密码子(UAA, UAG, UGA)时,翻译过程进入终止阶段。
单元步骤 1:释放因子的识别
I类释放因子(RF1识别UAA, UAG;RF2识别UAA, UGA)识别并结合到A位点的终止密码子上。这是一个特异性的结合反应,速率取决于终止密码子的类型和释放因子的浓度。在这个过程中主要涉及到释放因子RF1, 释放因子RF2这两个关键的复合物
70S_mRNA_pept-tRNA(P-site)_StopCodon(A-site) + RF1/RF2 <=> 70S_mRNA_pept-tRNA(P-site)_RF(A-site)单元步骤 2:肽链释放与核糖体解离
释放因子触发P位点上肽酰-tRNA的酯键水解,从而释放完整的多肽链。随后,在II类释放因子RF3和GTP的帮助下,RF1/RF2和脱酰化的tRNA从核糖体上解离。肽链的水解是另一个不可逆的化学步骤。RF3-GTP的作用类似于延伸因子,通过GTP水解促进其他因子的释放。
70S_mRNA_pept-tRNA(P-site)_RF(A-site) + H2O -> 70S_mRNA_deacyl-tRNA(P-site)_RF(A-site) + Peptide
70S_mRNA_deacyl-tRNA(P-site)_RF(A-site) + RF3-GTP -> 70S_mRNA_deacyl-tRNA(P-site) + RF(A-site) + RF3-GDP + Pi阶段四:核糖体再循环
终止后,70S核糖体、mRNA和脱酰化的tRNA仍然结合在一起。再循环过程将它们分离开,使核糖体亚基可以用于新一轮的翻译。
单元步骤:核糖体亚基的解离
这是翻译周期的最后一个关键步骤,确保了核糖体资源的有效利用。RRF和EF-G-GTP的协同作用是模型的重点。解离后的30S亚基会立即与IF3结合,防止其重新与50S亚基结合,从而可以开始新的起始过程。在这里最后一个阶段,核糖体再循环因子(RRF)与EF-G-GTP协同作用,将70S核糖体解离为30S和50S亚基,并释放mRNA和脱酰化的tRNA。
70S_mRNA_deacyl-tRNA + RRF + EF-G-GTP -> 30S + 50S + mRNA + deacyl-tRNA + RRF + EF-G-GDP + Pi翻译过程反应总结表
| 阶段 | 单元步骤 | 化学反应式 | 关键分子/复合物 | 建模考量 |
|---|---|---|---|---|
| 起始 | 30S小亚基与mRNA结合 | 30S + mRNA + IF1 + IF3 <=> 30S_mRNA_IF1_IF3 |
30S亚基, mRNA, IF1, IF3 | 可逆结合反应,需定义k_on和k_off。IF3防止50S亚基过早结合。 |
| 起始tRNA的装载 | 30S_mRNA_IF1_IF3 + fMet-tRNA^fMet_IF2-GTP <=> 30S_mRNA_fMet-tRNA^fMet_IF1_IF2-GTP_IF3 |
fMet-tRNA^fMet, IF2, GTP | 可逆结合,IF2-GTP确保起始tRNA的特异性。需考虑GTP浓度。 | |
| 50S大亚基结合与因子释放 | 30S..._IF2-GTP + 50S -> 70S_mRNA_fMet-tRNA^fMet + IF1 + IF2-GDP + IF3 + Pi |
50S亚基, GTP | GTP水解驱动的不可逆反应。正向速率远大于逆向。 | |
| 延伸 | 氨酰-tRNA进位 | 70S..._pept-tRNA(P) + aa-tRNA_EF-Tu-GTP -> 70S..._pept-tRNA(P)_aa-tRNA(A) + EF-Tu-GDP |
aa-tRNA, EF-Tu, GTP | 关键的保真性步骤。正确的密码子-反密码子配对会显著加速GTP水解。 |
| 肽键形成 | 70S..._pept-tRNA(P)_aa-tRNA(A) -> 70S..._deacyl-tRNA(P)_pept-aa-tRNA(A) |
70S核糖体 (肽酰转移酶) | 核心化学反应,速率快,可用k_cat描述。不直接消耗高能磷酸键。 |
|
| 转位 | 70S..._deacyl-tRNA(P)_pept-aa-tRNA(A) + EF-G-GTP -> 70S..._pept-aa-tRNA(P) + EF-G-GDP + Pi |
EF-G, GTP | GTP水解驱动的不可逆步骤,确保翻译单向进行。 | |
| 终止 | 释放因子识别 | 70S..._pept-tRNA(P)_StopCodon(A) + RF1/RF2 <=> 70S..._pept-tRNA(P)_RF(A) |
RF1, RF2 | 特异性结合反应,速率依赖于终止密码子类型和RF浓度。 |
| 肽链释放与因子解离 | 70S..._pept-tRNA(P)_RF(A) + H2O + RF3-GTP -> Peptide + 70S..._deacyl-tRNA(P) + RF + RF3-GDP + Pi |
RF3, GTP, H2O | 肽链水解不可逆。RF3-GTP促进其他因子释放,类似于延伸因子。 | |
| 再循环 | 核糖体亚基解离 | 70S_mRNA_deacyl-tRNA + RRF + EF-G-GTP -> 30S + 50S + mRNA + deacyl-tRNA + EF-G-GDP + Pi |
RRF, EF-G, GTP | GTP水解驱动的不可逆解离。RRF和EF-G协同作用,释放的30S亚基立即与IF3结合,准备新一轮起始。 |
- 单细胞视角下的微生物基因组代谢酶嵌入分析 - 2026年2月25日
- 基因组代谢酶层级嵌入 - 2026年2月23日
- 基因组功能吉布斯LDA主题建模 - 2026年2月23日

No responses yet